Itu Penghentian adalah fase terakhir dalam replikasi DNA. Ini didahului dengan inisiasi dan perpanjangan. Penghentian replikasi secara dini dapat mengakibatkan ekspresi protein yang diperpendek dan dengan demikian terjadi mutasi.
Apa pengakhirannya?

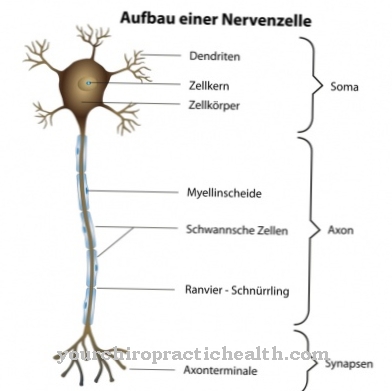
Selama replikasi atau reduplikasi, DNA pembawa informasi genetik diperbanyak dalam sel individu. Penggandaan terjadi menurut prinsip semi-konservatif dan biasanya mengarah pada penggandaan informasi genetik yang tepat. Replikasi dipicu selama fase sintesis, sebelum fase mitosis, dan dengan demikian terjadi sebelum inti sel membelah.
Pada awal replikasi, untai ganda DNA dipisahkan menjadi untai tunggal, di mana untai pelengkap baru terbentuk. Setiap untai DNA ditentukan oleh urutan basa untai yang berlawanan. Replikasi DNA terjadi dalam beberapa fase. Penghentian adalah fase replikasi ketiga dan karenanya terakhir. Penghentian diawali dengan inisiasi dan perpanjangan.
Istilah sinonim untuk mengungkapkan penghentian dalam konteks ini adalah sebutan Fase penghentian. Penghentian di sini berarti "membatalkan" atau "penghentian". Selama terminasi, untai mRNA yang baru terbentuk terlepas dari DNA sebenarnya. Pekerjaan DNA polimerase perlahan-lahan akan segera berakhir. Penghentian replikasi DNA tidak boleh disamakan dengan penghentian replikasi RNA.
Fungsi & tugas
Dalam fase inisiasi replikasi, terutama regulasi replikasi berlangsung. Titik awal replikasi ditentukan dan yang disebut priming terjadi. Setelah inisiasi, polimerisasi dimulai, selama fase perpanjangan dilewatkan. Enzim DNA polimerase memisahkan untaian komplementer DNA menjadi untaian tunggal dan membaca dasar untai tunggal satu demi satu. Dalam fase ini penggandaan semi-terputus berlangsung, yang mencakup fase pelapisan dasar lainnya.
Hanya setelah inisiasi dan perpanjangan barulah fase penghentian mengikuti dalam replikasi. Pengakhiran berbeda dari bentuk kehidupan ke bentuk kehidupan. Pada eukariota seperti manusia, DNA terstruktur dalam sebuah cincin. Ini juga mencakup urutan terminasi yang sesuai dengan dua urutan berbeda, yang masing-masing relevan untuk garpu replikasi.
Penghentian biasanya tidak dipicu oleh mekanisme khusus. Segera setelah dua cabang replikasi berjalan bersama atau DNA berakhir, replikasi secara otomatis diakhiri pada titik ini. Replikasi dihentikan dalam mekanisme otomatis.
Urutan penghentian adalah elemen kontrol. Mereka memastikan bahwa fase replikasi tiba di titik akhir tertentu dengan cara yang terkontrol meskipun kecepatan replikasi berbeda di dua garpu replikasi. Semua situs terminasi sesuai dengan situs pengikatan untuk protein Tus, "zat yang memanfaatkan terminal". Protein ini memblokir DnaB helikase replikatif dan dengan demikian menghentikan replikasi.
Pada eukariota, untaian cincin yang direplikasi tetap terhubung satu sama lain bahkan setelah replikasi. Sambungan sesuai dengan titik terminal. Hanya setelah pembelahan sel mereka dipisahkan oleh berbagai proses dan dengan demikian dapat dibagi. Koneksi yang tersisa sampai setelah pembelahan sel tampaknya memainkan peran dalam distribusi terkontrol.
Ada dua mekanisme utama yang berperan dalam pemisahan akhir cincin DNA. Enzim seperti tipe I dan tipe II topoisomerase terlibat dalam pemisahan. Akhirnya, protein tambahan mengenali kodon stop selama terminasi. Hal ini menyebabkan polipeptida terlepas dari ribosom, karena tidak ada t-RNA dengan antikodon yang sesuai untuk kodon stop yang tersedia. Ribosom akhirnya terurai menjadi dua subunitnya.
Penyakit & penyakit
Semua proses untuk duplikasi materi genetik dalam arti replikasi adalah rumit dan membutuhkan pengeluaran zat dan energi yang tinggi di dalam sel. Karena alasan ini, kesalahan replikasi spontan dapat dengan mudah terjadi. Jika genom berubah secara spontan atau diinduksi dari luar, kita berbicara tentang mutasi.
Kesalahan replikasi dapat mengakibatkan pangkalan hilang, dikaitkan dengan basa yang diubah, atau akibat dari pemasangan basa yang salah. Selain itu, penghapusan dan penyisipan nukleotida tunggal atau ganda di dalam dua untai DNA dapat menyebabkan kesalahan replikasi. Hal yang sama berlaku untuk dimer pirimidin, untai putus, dan kesalahan ikatan silang dalam untai DNA.
Mekanisme perbaikan terpisah tersedia jika terjadi kesalahan replikasi. Banyak dari kesalahan yang disebutkan diperbaiki sejauh mungkin dengan DNA polimerase. Akurasi replikasi relatif tinggi. Tingkat kesalahan hanya satu kesalahan per nukleotida, yang dapat dikaitkan dengan sistem kontrol yang berbeda.
Misalnya, mekanisme kontrol sel eukariotik dikenal sebagai peluruhan mRNA yang dimediasi nonsense, yang dapat mengenali kodon stop yang tidak diinginkan dalam mRNA dan dengan demikian mencegah protein yang diperpendek untuk menemukan ekspresi.
Kodon stop prematur dalam mRNA disebabkan oleh mutasi gen. Apa yang disebut mutasi nonsense atau alternatif dan penyambungan yang salah dapat mengakibatkan protein pendek yang dipengaruhi oleh kehilangan fungsional. Mekanisme kontrol tidak selalu dapat memperbaiki kesalahan.
Gangguan bawaan resesif autosomal β-thalassemia datang dalam tiga bentuk berbeda: yang pertama adalah thalassemia homozigot, penyakit serius yang disebabkan oleh mutasi nonsense Anda. Thalassemia heterozigot adalah penyakit yang lebih ringan di mana mutasi nonsense hanya ditemukan pada satu salinan gen β-globin. Melalui mekanisme peluruhan mRNA yang dimediasi nonsense, mRNA dari gen yang salah dapat terdegradasi sedemikian rupa sehingga hanya gen sehat yang diekspresikan.
Pada talasemia heterozigot dan dengan demikian bentuk penyakitnya sedang, mutasi nonsense berada pada mRNA ekson terakhir, sehingga mekanisme kontrol tidak diaktifkan. Untuk alasan ini, selain β-globin yang sehat, β-globin yang diperpendek juga terbentuk.Eritrosit dengan β-globin yang rusak akan binasa.
Contoh lain dari kegagalan mekanisme kontrol adalah distrofi muskular Duchenne, yang juga disebabkan oleh mutasi nonsense pada mRNA. Dalam hal ini, mekanisme kontrol memecah mRNA, tetapi dengan demikian menyebabkan hilangnya total protein distrofin.









.jpg)

.jpg)